המומחה הרפואי של המאמר

פרסומים חדשים
וירוס החיסוני האנושי (HIV)
סקירה אחרונה: 23.04.2024

כל תוכן iLive נבדק מבחינה רפואית או נבדק למעשה כדי להבטיח דיוק עובדתי רב ככל האפשר.
יש לנו קווים מנחים קפדניים המקור רק קישור לאתרים מדיה מכובד, מוסדות מחקר אקדמי, בכל עת אפשרי, עמיתים מבחינה רפואית מחקרים. שים לב שהמספרים בסוגריים ([1], [2] וכו ') הם קישורים הניתנים ללחיצה למחקרים אלה.
אם אתה סבור שתוכן כלשהו שלנו אינו מדויק, לא עדכני או מפוקפק אחרת, בחר אותו ולחץ על Ctrl + Enter.
תסמונת הכשל החיסוני שנרכשה הופרדה כמחלה מיוחדת בשנת 1981 בארצות הברית, כאשר במספר צעירים נגרמו מחלות קשות על ידי מיקרואורגניזמים שאינם פתוגניים או פתוגניים מעט עבור אנשים בריאים. בדיקת מצב החיסון של החולים חשפה ירידה חדה במספר הלימפוציטים בכלל ו- T-helpers בפרט. מצב זה נקרא איידס (אנגלית נרכשת חיסוני מחלת תסמונת - נרכש תסמונת אימונודיפייסי, או איידס). שיטת ההדבקה (מגע מיני, דרך הדם והכנותיו), הצביעה על האופי המזוהם של המחלה.
הסוכן הסיבתי של איידס התגלה בשנת 1983 באופן עצמאי על ידי הצרפתי ל. מונטנייר, שקרא לו LAV Lymphoadenopathy Associated Virus), כפי שהוא זיהה חולה עם לימפדנופתיה; ואת האמריקאי ר גאלו, אשר כינה את הנגיף HTLV-III (אנגלית האדם T-lymphotropic וירוס III): בעבר הוא נמצא וירוסים lymphotropic אני ו- II.
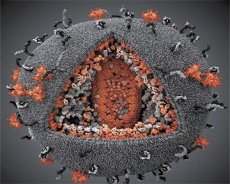
השוואה בין המאפיינים של נגיפי LAV ו- HTLV-III הראתה את זהותם, ולכן ב -1986 נקרא הנגיף HIV (HIV) (HIV), כדי למנוע בלבול. HIV הוא כדורית, קוטרו הוא 110 ננומטר. המעטפה של הנגיף יש צורה של polyhedron המורכב 12 מחומשים ו 20 משושים. מולקולה של gpl20 glycylylated חלבון ממוקם במרכז ופינות של כל משושה (מספר 120 פירושו משקל מולקולרי של חלבון קילו-דלונים). סך של 72 מולקולות gpl20 ממוקמים על פני השטח של virion, שכל אחד מהם קשורה חלבון intramembrane gp41. חלבונים אלה יחד עם שכבת השומנים כפולה טופס supercapsid (ממברנה) של virion.
חלבונים gpl20 ו gp41 נוצרים כתוצאה של חיתוך פרוטאז התא של מבשר חלבון Env. חלבון gp41 יוצר את "חתיך" של עמוד השדרה, להיות מקושר על ידי תחום cytoplasmic עם p17MA חלבון מטריקס מיד מתחת לממברנה. מולקולות p17, אינטראקציה עם התבגרות של virion, טופס icosahedron מתחת לקליפה.
בחלק המרכזי של virion, חלבון p24 יוצר קפסיד חרוטי. החלק הצר של הקפסיד עם השתתפות של חלבון pb מחובר עם המעטפה של virion. בתוך הקפסיד, יש שתי מולקולות זהות של RNA גנומי ויראלי. הם קשרו את הקצוות 5 'שלהם p7NC nucleocapsid חלבון. חלבון זה הוא מעניין, כי יש לו שתי שאריות חומצת אמינו (מוטיב), ציסטאין-עשיר אטום היסטידין ו המכיל Zn, - הם נקראים "אצבע אבץ", כשהם מצלמים את המולקולה של RNA הגנומי לצורך שילובם virions יצר. הקפסיד מכיל גם שלושה אנזימים. Reversase (RT), או pol-complex, כולל transcriptase הפוכה, RNA-Ase H, ו- DNA תלויי פולימראז DNA. Revertase קיים כמו heterodimer p66 / p51. Protease (PR) - pI, מתחיל ומבין את תהליך ההתבגרות virion. אינטגרציה (IN) - p31, או endonuclease, מבטיחה הכללת דנ"א פרופירי בגנום של התא המארח. הקפסיד מכיל גם מולקולת זרע RNA (tRNAl "3).
הגן RNA בתא מומרת על ידי transcriptase לאחור לתוך הגנום DNA (DNA-provirus) בהיקף של 9283 זוגות נוקליאוטידים. זה מוגבל לשמאל ולימין על ידי מה שמכונה סוף סוף חוזר, או LTR (אנגלית לחזור מסוף ארוך): S'-LTR - שמאל Z'-LTR - בצד ימין. LTR מכילים 638 זוגות נוקליאוטידים.
גנום ה- HIV מורכב מ -9 גנים, שחלקם חופפים (יש מספר מסגרות קריאה) ויש להם מבנה אקסונין. הם שולטים בסינתזה של 9 חלבונים מבניים ו -6 רגולטוריים.
ערך ה- LTR של הגנום הנגיפי הוא שהם מכילים את האלמנטים הרגולטוריים הבאים השולטים בתפקודו:
- אות תעתיק (אזור האמרגן);
- האות להוספת פולי-A;
- לכידת האות;
- שילוב אותות;
- איתות רגולטורי חיובי (TAR עבור חלבון TAT);
- אלמנט של רגולציה שלילית (NRE עבור חלבון NEF);
- האתר המצורף של RNA זרע (tRNA ™ 3) עבור סינתזה של שרשרת מינוס של ה- DNA בסוף 3 '; האות ב 5'-end של LTR, המשמשת תחל עבור סינתזה של פלוס- strand של ה- DNA.
בנוסף, LTR מכיל אלמנטים המעורבים הרגולציה של שחבור mRNA, אריזה מולקולות vRNA לתוך capsid (אלמנט Psi). לבסוף, כאשר תעתיק הגנום ב mRNAs ארוך, שני אותות נוצרים עבור חלבון REV, אשר לעבור סינתזה חלבון: CAR עבור חלבונים רגולטוריים CRS עבור חלבונים מבניים. אם חלבון REV נקשר לרכב, חלבונים מבניים מסונתזים; אם הוא נעדר, רק חלבונים רגולטוריים מסונתזים.
בויסות הגנום של הנגיף, הגנים המסדירים והחלבונים שלהם ממלאים תפקיד חשוב במיוחד:
- חלבון TAT שמבצע בקרה חיובית על ההתרבות של הנגיף ופועל דרך האתר TAR הרגולציה;
- חלבונים NEV ו VPU, ביצוע שליטה שלילית של רבייה באמצעות אתר NRE;
- חלבון REV, ביצוע שליטה שלילי חיובי. חלבון REV שולט על העבודה של גנים gag, פול, env ומבצע רגולציה שלילית של שחבור.
לפיכך, הרבייה של HIV נמצאת תחת שליטה משולשת - חיובי, שלילי וחיובי שלילי.
חלבון ה- VIF קובע את infectivity של וירוס מסונתז החדש. זה קשור אל p24 חלבון capsid והוא נוכח virion בסכום של 60 מולקולות. חלבון NEF מיוצג ב virion על ידי מספר קטן של מולקולות (5-10), אולי קשור המעטפה.
חלבון VPR מעכב את מחזור התא בשלב G2, משתתף בהובלת מתחמי preintegration לתוך גרעין התא, מפעיל כמה גנים ויראליים וסלולריים, מגביר את היעילות של שכפול ויראלי מונוציטים מקרופאגים. המיקום של חלבונים VPR, TAT, REV, VPU ב virion לא הוקמה.
בנוסף חלבונים משלה, הרכב של קרום virion עשוי לכלול כמה חלבונים של התא המארח. חלבונים VPU ו VPR מעורבים הרגולציה של הרבייה וירוס.
גרסאות אנטיגניות של נגיף האיידס (HIV)
וירוס החיסונים האנושי (HIV) הוא משתנה מאוד. אפילו מן האורגניזם של מטופל אחד, ניתן לבודד את זני הנגיף השונים באופן משמעותי בתכונות האנטיגניות. השתנות זו מקודמת על ידי הרס אינטנסיבי של תאי CD4 + ותגובת נוגדנים חזקה לדלקת ב- HIV. חולים ממערב אפריקה יש צורה חדשה של HIV, קרוב ביולוגית ל- HIV-1, אבל נבדלת באופן אימונולוגי ממנו, HIV-2. ההומולוגיה של המבנה העיקרי של הגנום של וירוסים אלה הוא 42%. DNA-provirus HIV-2 מכיל 9671 bp, ו- LTR - 854 bp. HIV-2 היה מבודד לאחר מכן באזורים אחרים של העולם. אין חסינות צולבת בין HIV-1 ו- HIV-2. שתי צורות גדולות של HIV-1 ידועים: O (Outlier) ו- M (מייג 'ור), האחרונים מחולקים 10 תת סוגים (AJ). ברוסיה, 8 subtypes להפיץ (AH).
 [
[מנגנון האינטראקציה של HIV עם התא
לאחר שחדר את הגוף, הנגיף הראשון התקפות תאים המכילים קולטן CD4 מסוים. קולטן זה יש מספר רב של T- עוזרים, פחות - מקרופאגים מונוציטים, במיוחד את ה- T רגישים ל-וירוסים.
וירוס החיסוני האנושי (HIV) מזהה קולטני CD4 עם חלבון gpl20 שלה. תהליך האינטראקציה של HIV עם התמורה התא על פי התוכנית הבאה: קולטן בתיווך ספיחה -> גובלת fovea -> גדילה שלפוחית -> lysosome. בתוך זה, קרום virion מתמזגת עם קרום lysosome, ואת nucleocapsid, משוחרר מן supercapsid, נכנס הציטופלסמה; בדרך לגרעין הוא נהרס, ו- RNA גנומי ומרכיבי הליבה הקשורים משוחררים. ואז transcriptase הפוך מסנתז את שרשרת מינוס של ה- DNA על רנ"א virion, אז RNA-Ase H הורס את RNA virion, ואת פולימראז DNA ויראלי מסנתזת פלוס- strand של ה- DNA. על הקצוות של DNA-provirus, 5'-LTR ו 3'-LTR נוצרים. DNA- provirus יכול להיות בגרעין במשך זמן מה בצורה לא פעילה, אבל במוקדם או במאוחר זה משתלב בעזרת integrase שלה לתוך הכרומוזום של תא המטרה. בו, provirus הוא במצב לא פעיל עד T- לימפוציט זה מופעל על ידי אנטיגנים חיידקים או תאים אחרים immunocompetent. הפעלת תעתיק של דנ"א סלולר מוסדר על ידי גורם גרעיני מיוחד (NF-kB). זהו חלבון מחייב DNA והוא מיוצר כמויות גדולות במהלך ההפעלה והתפשטות של לימפוציטים T ומונוציטים. חלבון זה נקשר לרצפים ספציפיים של DNA תאיים ורצפים דומים של LTR DNA-LTR וגורם שעתוק של ה- DNA הסלולרי ו- DNA-provirus. על ידי יצירת שעתוק של ה- DNA-provirus, הוא מעביר את הנגיף ממצב לא פעיל לדלקת פעילה, ולפיכך זיהום מתמשך, לתוך זיהום פרודוקטיבי. להישאר provirus במצב לא פעיל יכול להימשך זמן רב מאוד. הפעלת הנגיף היא רגע קריטי באינטראקציה עם התא.
מרגע כניסת הנגיף לתא, מתחילה תקופה של הידבקות ב- HIV, וירוס שיכול להימשך 10 שנים או יותר; ומאז ההפעלה של הנגיף מתחיל המחלה - איידס. בעזרת הגנים הרגולטוריים שלהם ומוצריהם, הווירוס מתחיל להתרבות באופן פעיל. חלבון TAT יכול להגדיל את שיעור הרבייה של הנגיף 1000 פעמים. תעתיק הנגיף הוא מורכב. הוא כולל את היווצרות של שניהם אורך מלא mRNAs subgenomic, שחבור של mRNA, סינתזה נוספת של חלבונים מבניים ורגולטוריים.
סינתזה של חלבונים מבניים מתרחשת כדלקמן. ראשית, מבשר polyprotein Pr55Gag הוא מסונתז (חלבון עם מסה של 55 kD). הוא מכיל ארבעה תחומים עיקריים: מטריצה (MA), הקופסית (CA), nucleocapsid (NC) ו PE תחום, מתוכם לגרום חיתוך פרוטאז ויראלי Pr55Gag (זה samovyrezaetsya מעוד מבשר חלבון - Gag-Pol) נוצרות בהתאמה P17 חלבונים מבניים , p24, p7 ו- pb. היווצרות של polyprotein Pr55Gag הוא התנאי העיקרי להיווצרות של חלקיקים נגיפיים. זה חלבון זה קובע את תוכנית morphogenesis של virion. היא כוללת צעדי תחבורת polyprotein ברצף Gag קרום הפלזמה והאינטראקציה שלו עם אינטראקציות חלבון-חלבון היווצרות חלקיקי וירוס הבוסר שלה. Pr55Gag הוא מסונתז על polyribosomes חינם; מולקולות חלבון מועברות אל הקרום, המעוגן לפי האזורים הידרופובי שלהם. התפקיד העיקרי ביצירת קונפורמציה מקומית של חלבון GAG הוא שיחק על ידי תחום CA. מתג NC-תחום מספק (באמצעות "אצבעות אבץ") 2 מולקולה של RNA הגנומי לתוך החלקיק הנגיפי להרכיב. מולקולת polyprotein הוא dimerized הראשון בשל האינטראקציה של תחומים מטריקס. ואז הדימרים לשלב hexameric (6 יחידות) מתחמי ידי האינטראקציה תחומים CA ו- NC. לבסוף, hexamers, משטחי הצד חיבור מהווים virions בשלה צורה כדורית, בתוך אשר מכילים שנתפסו RNA הגנומי ויראלי-תחום NC.
עוד מבשר חלבון Prl60Gag-Pol (החלבון עם מ. מ 160 KDA) הוא מסונתז כתוצאה הסטה את מסגרת הקריאה של הריבוזום במהלך הסופית- Z'התרגום של הגן איסור הפרסום לאזור מיד לפני קידוד חלבון באזור RB. פוליפרוטין Gag-Pol זה מכיל רצף שלם של חלבון Gag (1 - 423 חומצות אמינו) ו רצפים פול, הכוללים יחסי ציבור, RT ו IN תחומים. מולקולות של פוליפרוטין Gag-Pol מסונתזים גם על polyriosomes חופשי מועבר קרום הפלזמה. Poliprotein Prl60Gagpol מכיל את כל האתרים של אינטראקציות intermolecular הטמון poliprotein גאג ואת אתרי מחייב הממברנה. לכן, המולקולות של פתיל Gag-Pol polyprotein עם הממברנה, יחד עם Gag-מולקולות כוללות virions להרכיב, אשר עלול לגרום לתהליך התבגרות פרוטאז virion פעיל מתחיל. ה- HIV-1 פרוטאז פעיל מאוד רק בצורה של דימר, ולכן, עבור כריתה עצמית של Prl60Gag-Pol, dimerization של מולקולות אלה נדרשת. התבגרות של virion היא כי פרוטאז פעיל פרוצים חתכים prl60Gag-Pol ו Gag55 לאתרים לזיהוי; חלבונים p17, p24, p7, p6, revertase, integrase נוצרים ואת הקשר שלהם במבנה ויראלי מתרחש.
חלבון מעטפה מסונתז על ריבוזומים הקשורים ממברנות של reticulum endoplasmic, glycosylated ואז הוא חתך פרוטאז הסלולר כדי gp120 ו- gp41 ומועבר אל פני התא. במקרה זה, gp41 מחלחל את הממברנה וקושר לתחומי המטריצה של מולקולת חלבון גאג הקשורים המשטח הפנימי של הממברנה. יחסים אלה נמשכים ב virion בוגרת.
לכן, הרכבה של חלקיקי הנגיף הוא אגרגציה של חלבונים מבשר מולקולות RNA הקשורות מופעל על הממברנה של התא המארח, היווצרות של virions בשלה שחרורם באמצעות הנצה מתא השטח. כאשר ניצני, virion מקיפה את עצמה עם קרום התא, שבו gp41 ו gp120 מולקולות מוטבעים. במהלך נביטה או אולי לאחר שחרורו של התבגרות virion מתרחש, אשר מתבצע באמצעות הפרוטאז נגיפי חלבונים מבשרים Pr55Gag חיתוך פרוטאוליטי ו Prl60Gag-Pol וירוס להתבגר חלבוני הקשר שלהם אל מתחמים מבניים ספציפיים. תפקיד מוביל בתהליכים של מורפוגנזה וירוס הוא שיחק על ידי מבשר polyprotein Pr55Gag, אשר מארגן ומרכיב virion בוגרת; תהליך ההתבגרות שלה הושלם על ידי פרוטאז ויראלי מסוים.
גורם לחוסר חיסוני
אחד הגורמים העיקריים לחוסר חיסוני בזיהום איידס הוא מוות המוני של T-helpers. זה קורה בגלל האירועים הבאים. ראשית, T- עוזר וירוסים נגועים על ידי הנגיף למות עקב אפופטוזיס. הוא האמין כי חולים עם שכפול נגיף האיידס, אפופטוזיס ירידה במספר תאי T-helper מחוברים ביניהם. שנית, תאי T-הרוצח לזהות ולהשמיד תאי T נגועים בוירוס או הנושא את מתרכובת gpl20, כמו גם נגוע וירוס נגוע בנגיף תאי T-helper, המהווים symplasts (syncytia) מורכב מכמה עשרות תאים (חלק הם מתים כתוצאה מכפל הווירוסים בהם). עקב ההרס של מספר רב של תאי T-helper מתרחש ביטוי הקולטן קרום ירידה לימפוציטים מסוג B ל IL-2, סינתזה מופרעת של אינטרלויקנים שונים (גורמי גדילה והתמיינות של לימפוציטים מסוג B -. IL-4, IL-5, IL-6, ואחרים) כתוצאה מכך הפונקציה של מערכת T-killer מופרת. דיכוי של פעילות של מערכות השלמה מקרופאג מתרחשת. מקרופאגים ומונוציטים שנדבקו בנגיף ארוך למות, אבל הם לא מסוגלים להסיר את הווירוס מהגוף. לבסוף, בשל הדמיון המבני ואת אנטיגני עם קולטנים gpl20 כמה תאי האפיתל של האורגניזם (כולל trophoblasts קולטן בתיווך העברת השתלת HIV) הוא מסונתז antiretseptornyh נוגדנים עם קשת רחבה של פעולה. נוגדנים כאלה יכולים לחסום קולטנים סלולריים שונים ולסבך את מהלך המחלה עם הפרעות אוטואימוניות. התוצאה של זיהום HIV היא התבוסה של כל החלקים העיקריים של המערכת החיסונית. חולים אלה הופכים לחסרי הגנה כנגד מגוון רחב של מיקרואורגניזמים. זה מוביל להתפתחות זיהומים אופורטוניסטיים ומחלות ניאופלסטיות. בחולים עם זיהום ב- HIV, לפחות שלושה סוגים של סרטן היו בסיכון מוגבר: סרקומה של Kaposi; קרצינומה (כולל סרטן העור); לימפומה מסוג B הנובעת מהתנוונות ממאירה של לימפוציטים מסוג B. עם זאת, HIV לא רק לימפוציטים, אלא גם נוירוטרופי. היא חודר בתאי CNS (אסטרוציטים) או על ידי אנדוציטוזה קולטן בתיווך ו phagocytosis של האסטרוציטים עם lymphoblasts הנגוע בנגיף. בשנת האינטראקציה של הנגיף עם האסטרוציטים נוצרים גם symplasts תורמת להתפשטות ערוצי אינטר הפתוגן. מקרופאגים ומונוציטים, הווירוס עלול להימשך זמן רב, כך הם משרתים כמאגר והמפיצים שלה בגוף, יכולת לחדור לתוך כל הרקמות. מקרופאגים נגועים יש תפקיד מרכזי הגירה של HIV במערכת העצבים המרכזית התבוסה שלה. בשנת 10% מהחולים עם תסמונות קליניות ראשוניות הקשורות למערכת העצבים המרכזית ואת המוצג בצורה של דמנציה (שיטיון). לכן, עבור אנשים מושפעים הידבקות ב- HIV מאופיין 3 קבוצות של מחלות - זיהומים אופורטוניסטיים, מחלות גידול CNS.
אפידמיולוגיה של הידבקות ב- HIV
המקור של זיהום HIV הוא רק אדם - חולה או וירוס נושא. וירוס החיסונים האנושי (HIV) נמצא בדם, הזרע, נוזל צוואר הרחם; באמהות מניקות - בחלב אם. זיהום מתרחש מינית, דרך הדם ותרופותיו, וכן מאם לילד לפני הלידה, במהלך ואחרי הלידה. מקרים של זיהום על ידי הנגיף באמצעות מזון, משקאות באמצעות עקיצות חרקים אינם ידועים.
התמכרות לסמים תורמת להתפשטות האיידס. זיהום HIV גדל מדי שנה. לדברי ארגון הבריאות העולמי, מ 1980 עד 2000, 58 מיליון בני אדם היו נגועים ב- HIV. רק בשנת 2000, 5.3 מיליון בני אדם נדבקו בעולם, ו -3 מיליון בני אדם מתו מאיידס. ברוסיה, החל ב -1 בינואר 2004, היו רשומים 264,000 אנשים עם HIV. מחצית מהנגועים ב- HIV מתים תוך 11-12 שנים של זיהום. בתחילת 2004, מתוך כל 100,000 אזרחי רוסיה, כ 180 חיו עם אבחנה של "הידבקות ב- HIV". זה צפוי כי ברמה זו של שכיחות, המספר הכולל של אנשים נגועים ב- HIV ברוסיה עד 2012 יהיה 2.5-3 מיליון בני אדם. המורכבות של המאבק נגד זיהום HIV תלוי במספר סיבות: ראשית, אין שיטות יעילות של הטיפול שלה ומניעה ספציפית; שנית, תקופת הדגירה לזיהום HIV יכולה לעלות על 10 שנים. משך הזמן תלוי ברגע ההפעלה של הלימפוציטים T ו- DNA-provirus הכלולים בכרומוזום. עדיין לא ברור אם כל נגיף נגוע באיידס הוא נדון או צפוי להיות וירוס לטווח ארוך ללא מחלה (אשר נראה סביר). לבסוף, ישנם מספר וירוסים של חיסונים אנושיים (HIV-1, HIV-2), הבדלים אנטיגניים ביניהם למנוע את היווצרות של חסינות צלב. גילוי של וירוס חיסוני של קופים (SIV) לשפוך אור על מקורו של HIV. ה- SIO לארגון הגנום דומה ל- HIV, אך הוא שונה משמעותית ברצף הנוקליאוטידים. HIV-2 נמצא במצב ביניים בין HIV-1 ל- SIV, ורצף הנוקליאוטידים היה קרוב יותר ל- SIV. בהקשר זה, VM Zhdanov הציע כי הנגיפים HIV-1, HIV-2 ו SIV שמקורה אב קדמון משותף. זה אפשרי, על פי ר 'גאלו, כי אחד SIVs איכשהו נכנס לגוף האדם, שם הוא עבר מספר מוטציות, אשר הביא HIV-1, HIV-2 וצורות אחרות של זה.
סימפטומים של הידבקות ב- HIV
הווירוס של חוסר החיסון האנושי מאופיין בתכונות מסוימות, שבהן הפתוגנזה של המחלה תלויה במידה רבה. לנגיף יש שיעור רבייה גבוה מאוד, שנקבע על ידי האלמנטים הרגולטוריים שלו (5,000 virions מסונתזים בשלב הפעיל תוך 5 דקות). בשל נוכחותו של חלבון היתוך (gp41), הנגיף גורם להיווצרות של מבנים syncytial נרחב עקב היתוך של נגוע T- עוזרים נגוע, וכתוצאה מכך המוות המונית שלהם. מולקולה גדולה מולקולות gpl20 להפיץ באופן חופשי בדם להיקשר קולטנים של T- עוזרי T- נגוע, וכתוצאה מכך הם מוכרים גם נהרס על ידי רוצחי T. הווירוס יכול להתפשט דרך התעלות הבין תאיות מהתא אל התא, במקרה זה הוא הופך להיות פחות מוכנה לנוגדנים.
קריטריונים קליניים לזיהום ב- HIV
HIV למבוגרים לקבוע אם יש להם לפחות שני סימפטומים חמורים בשילוב עם סימפטום אחד לפחות של קטין בהיעדר גורמים ידועים אחרים של חסר חיסוני (סרטן, כשל חיסוני מולד, רעב חמור, וכן הלאה. פ). תסמינים חמורים כוללים:
- ירידה במשקל של 10% או יותר;
- קדחת ממושכת, לסירוגין או מתמשכת;
- שלשולים כרוניים.
תסמינים קלים: שיעול מתמשך, דרמטיטיס כללית, הרפס זוסטר חוזר ונשנה, קנדידיזיס של חלל הפה והלוע, הרפס סימפלקס כרוני, לימפדנופתיה כללית. אבחנה של איידס נעשה עם נוכחות של רק סרקומה של קפוסי, דלקת קרום המוח קריפטוקוקלית, דלקת ריאות pneumocystis. התמונה הקלינית של המחלה מושפעת מזיהום אופורטוניסטי.
שיטות טיפוח של וירוס החיסוני האנושי (HIV)
HIV-1 ו- HIV-2 ניתן לתרבית בתאי שיבוט אחד בלבד של לימפוציטים TCB4 - H9, המתקבלים מלימפוציטים TCV4 לוקמיה. עבור אותן מטרות, תרבויות monolayer של תאים astrocyte יכול לשמש גם, שבו HIV-1 מכפיל היטב. מבעלי חיים לשימפנזים רגישים ל- HIV-1.
ההתנגדות של הנגיף בסביבה החיצונית נמוכה. הוא מת תחת השפעת אור השמש והקרנה UV, נהרס ב 80 מעלות צלזיוס למשך 30 דקות, כאשר מטופלים עם חיטוי נפוץ - במשך 20-30 דקות. כדי לחטא את החומר המכיל וירוסים, יש צורך להשתמש חיטוי mycobactricidal, שכן הם יעילים נגד מיקרואורגניזמים עם ההתנגדות הגבוהה ביותר.
אבחון מעבדה של הידבקות ב- HIV
הדרך העיקרית לאבחן את הנגיף וזיהום ה- HIV הוא החיסון האינסופי. עם זאת, בשל העובדה כי יש gpl20 מבני דמיון אנטיגני לקולטנים של תאים אנושיים מסוימים, כוללים קולטנים מבצעות הובלה של אימונוגלובולינים דרך תאי האפיתל של הריריות בגוף עשוי להופיע נוגדנים הקשורים נוגדנים נגד gpl20. במקרה זה, יכולות להיות תוצאות חיוביות שגויות של ה- IFM. לכן, כל תגובה חיובית של סרה של הנחקר נתונים לניתוח נוסף על ידי השיטה אימונובלוט, או המערבי סופג. שיטה זו מתבססת על זיהוי הנוגדנים הנלמדים לאחר הפרדה אלקטרופורטית ובדיקה עוקבת עם נוגדן אנטי-וירוס שכותרתו. השיטה הווירולוגית אינה מועילה בשל המורכבות של תרבות הנגיף. שיבוט של לימפוציטים H9 משמש כדי להשיג אנטיגנים ויראליים - המרכיבים הדרושים של מערכות בדיקה אבחון. שיטת CDR מאפשרת לזהות את הנגיף כבר בשלב מוקדם של וירמיה.
טיפול בזיהום ב- HIV
יש צורך למצוא או לסנתז תרופות המעכבות את הפעילות של transcriptase הפוך (revertase) או פרוטאז ויראלי. הם היו מונעים היווצרות של DNA- provirus ו (או) לעכב את הכפל תאיים של הנגיף. האסטרטגיה החדשה של הטיפול של נשאי HIV מבוססת על העיקרון של שימוש בשילוב של תרופות המעכבות את הפרוטאז הנגיפי (אחת התרופות) ו revergazu (2 סמים שונים), - בשילוב טיפול (טריפל). ברוסיה, לטיפול אנשים נגועים ב- HIV מומלץ לשימוש בשילוב של שתי תרופות מקומיות: Crixivan Phosphazide ובאופן ספציפי לעכב שעתוק של HIV בשלבים מוקדמים ומאוחרים של הרבייה, במיוחד עם פעילות מופחתת של AZT.
הבעיה של מניעה ספציפית היא הצורך ליצור חיסון שיבטיח היווצרות של חסינות יעילה בתיווך תא המבוססת על לימפוציטים ציטוטוקסיים ספציפיים לוירוס ללא כל ייצור משמעותי של נוגדנים. חסינות כזו מסופקת על ידי Thl-helpers. יתכן כי נוגדנים, כולל נטרול וירוסים, אינם רק לא יעיל בדיכוי זיהום HIV, אך ברמה גבוהה הם מדכאים חסינות בתיווך התא. לכן, החיסון נגד HIV צריך לעמוד בראש ובראשונה שתי דרישות בסיסיות: א) להיות בטוח לחלוטין ב) לעורר את פעילותם של לימפוציטים T-cytotoxic. יעילותם של גרסאות שונות של חיסונים המתקבלים מוירוסים שנהרגו (מומתים) ומאנטיגנים אינדיבידואלים בעלי תכונות הגנה גבוהות נבדקת. אנטיגנים כאלה יכולים להיות מבודדים מן virions עצמם, או מסונתז כימית. חיסון המבוסס על שיטות הנדסה גנטית מוצע. זהו וירוס חיסון רקומביננטי הנושא גנים של HIV האחראים לסינתזה של אנטיגנים בעלי תכונות חיסוניות חזקות. ההחלטה על האפקטיביות של חיסונים אלה לוקח זמן רב בגלל משך זמן ארוך של תקופת הדגירה של זיהום HIV ואת השונות הגבוהה של הפתוגן. יצירת חיסון יעיל נגד HIV הוא בעיה בסיסית דחופה.