המומחה הרפואי של המאמר

פרסומים חדשים
בלוטת יותרת המוח
סקירה אחרונה: 04.07.2025

כל תוכן iLive נבדק מבחינה רפואית או נבדק למעשה כדי להבטיח דיוק עובדתי רב ככל האפשר.
יש לנו קווים מנחים קפדניים המקור רק קישור לאתרים מדיה מכובד, מוסדות מחקר אקדמי, בכל עת אפשרי, עמיתים מבחינה רפואית מחקרים. שים לב שהמספרים בסוגריים ([1], [2] וכו ') הם קישורים הניתנים ללחיצה למחקרים אלה.
אם אתה סבור שתוכן כלשהו שלנו אינו מדויק, לא עדכני או מפוקפק אחרת, בחר אותו ולחץ על Ctrl + Enter.
בלוטת יותרת המוח (hypophysis, s.glandula pituitaria) ממוקמת בגומה ההיפופיזה של העצם הטורקית (sella turcica) בעצם הספנואיד ומופרדת מחלל הגולגולת על ידי תהליך של הדורה מאטר (dura mater) במוח, היוצר את הסרעפת של העצם הטורקית. דרך פתח בסרעפת זו, בלוטת יותרת המוח מחוברת לאינפונדיבולום (infundibulum) של ההיפותלמוס (diencephalon). הגודל הרוחבי של בלוטת יותרת המוח הוא 10-17 מ"מ, הקדמי-אחורי הוא 5-15 מ"מ, האנכי הוא 5-10 מ"מ. מסת בלוטת יותרת המוח אצל גברים היא כ-0.5 גרם, אצל נשים - 0.6 גרם. בלוטת יותרת המוח מכוסה חיצונית בקפסולה.
בהתאם להתפתחות בלוטת יותרת המוח משני יסודות שונים, נבדלות שתי אונות באיבר - הקדמית והאחורית. האדנוהיפופיזה, או האונה הקדמית (adenohypophysis, s.lobus anterior), גדולה יותר, ומהווה 70-80% מהמסה הכוללת של בלוטת יותרת המוח. היא צפופה יותר מהאונה האחורית. באונה הקדמית, מבחין חלק דיסטלי (pars distalis), התופס את החלק הקדמי של גומת יותרת המוח, חלק ביניים (pars intermedia), הממוקם על הגבול עם האונה האחורית, וחלק פקעתי (pars tuberalis), העולה כלפי מעלה ומתחבר לאינפונדיבולום של ההיפותלמוס. בשל שפע כלי הדם, לאונה הקדמית יש צבע צהוב חיוור עם גוון אדמדם. הפרנכימה של בלוטת יותרת המוח הקדמית מיוצגת על ידי מספר סוגים של תאים בלוטיים, שבין גדיליהם ממוקמים נימים סינוסואידליים. מחצית (50%) מתאי האדנוהיפופיזה הם אדנוציטים כרומופיליים, בעלי גרגירים דקים בציטופלזמה שלהם, הנצבעים היטב במלחי כרום. אלו הם אדנוציטים אצידופיליים (40% מכלל תאי האדנוהיפופיזה) ואדנוציטים בזופיליים (10%). אדנוציטים בזופיליים כוללים אנדוקרינוציטים גונדוטרופיים, קורטיקוטרופיים ותירוטרופיים. אדנוציטים כרומופוביים הם קטנים, בעלי גרעין גדול וכמות קטנה של ציטופלזמה. תאים אלה נחשבים לתאים קודמנים של אדנוציטים כרומופיליים. 50% הנותרים של תאי האדנוהיפופיזה הם אדנוציטים כרומופוביים.
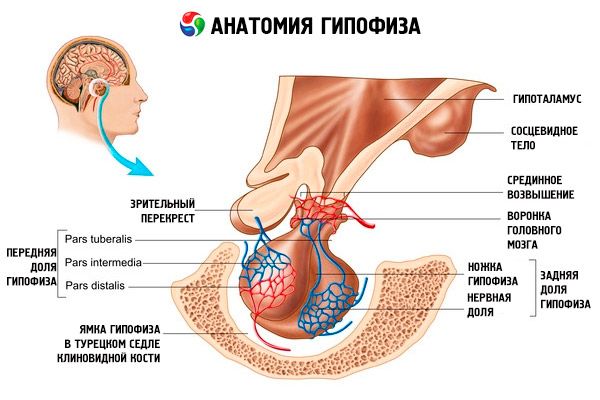
הנוירו-היפופיזה, או האונה האחורית (neurohypophysis, s.lobus posterior), מורכבת מהאונה העצבית (lobus nervosus), הממוקמת בחלק האחורי של גומת בלוטת יותרת המוח, ומהמשפך (infundibulum), הממוקם מאחורי החלק הפקעתי של האדנו-היפופיזה. האונה האחורית של בלוטת יותרת המוח נוצרת על ידי תאי נוירוגליה (תאי יותרת המוח), סיבי עצב העוברים מגרעיני ההפרשה הנוירו-עצבית של ההיפותלמוס לנוירו-היפופיזה, וכדוריות נוירו-הפרשה.
בלוטת יותרת המוח, באמצעות סיבי עצב (מסלולים) וכלי דם, מחוברת באופן פונקציונלי להיפותלמוס של ה-diencephalon, המווסת את פעילות בלוטת יותרת המוח. בלוטת יותרת המוח וההיפותלמוס, יחד עם הקשרים הנוירואנדוקריניים, כלי הדם והעצביים שלהם, נחשבים בדרך כלל למערכת ההיפותלמוס-יותרת המוח.
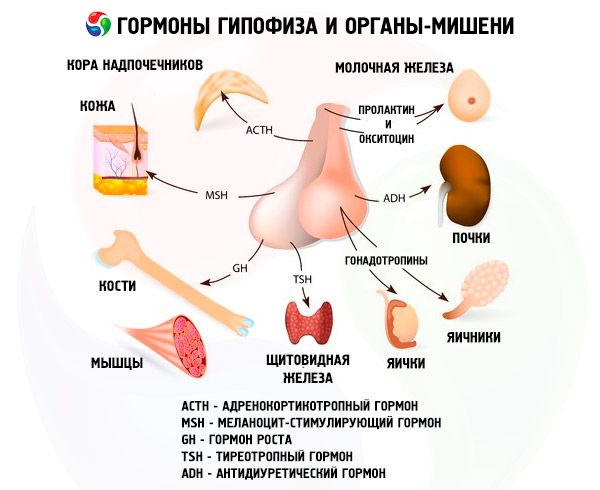
הורמונים של בלוטות יותרת המוח הקדמית והאחורית משפיעים על תפקודים רבים בגוף, בעיקר דרך בלוטות אנדוקריניות אחרות. בבלוטת יותרת המוח הקדמית, אדנוציטים אצידופיליים (תאי אלפא) מייצרים הורמון סומטוטרופי (הורמון גדילה), אשר משתתף בוויסות תהליכי גדילה והתפתחות באורגניזמים צעירים. אנדוקרינוציטים קורטיקוטרופיים מפרישים הורמון אדרנוקורטיקוטרופי (ACTH), אשר מגרה את הפרשת הורמוני הסטרואידים על ידי בלוטות יותרת הכליה. אנדוקרינוציטים תירוטרופיים מפרישים הורמון תירוטרופי (TSH), אשר משפיע על התפתחות בלוטת התריס ומפעיל את ייצור ההורמונים שלה. הורמונים גונדוטרופיים: מגרה זקיק (FSH), לוטיניזציה (LH) ופרולקטין - משפיעים על ההתבגרות המינית של הגוף, מווסתים ומגרים את התפתחות הזקיקים בשחלה, ביוץ, צמיחת בלוטות החלב וייצור חלב אצל נשים, ואת תהליך הזרע אצל גברים. הורמונים אלה מיוצרים על ידי אדנוציטים בזופיליים (תאי בטא). כאן מופרשים גם גורמים ליפוטרופיים של בלוטת יותרת המוח, אשר משפיעים על גיוס וניצול שומנים בגוף. בחלק הביניים של האונה הקדמית נוצר הורמון מגרה מלנוציטים, אשר שולט על היווצרות פיגמנטים - מלנינים - בגוף.
תאים נוירו-פרשניים של הגרעינים הסופראופטיים והפרה-חדריים בהיפותלמוס מייצרים וזופרסין ואוקסיטוצין. הורמונים אלה מועברים לתאי בלוטת יותרת המוח האחורית דרך אקסונים המרכיבים את מערכת ההיפותלמוס-יותרת המוח. מבלוטת יותרת המוח האחורית, חומרים אלה נכנסים לדם. להורמון וזופרסין יש השפעה מכווצת כלי דם ואנטי-משתנת, שבגללה הוא נקרא גם הורמון אנטי-משתן (ADH). לאוקסיטוצין יש השפעה מעוררת על התכווצות שרירי הרחם, מגביר את הפרשת החלב על ידי בלוטת החלב המניקה, מעכב את התפתחות ותפקוד הגופיף הצהוב, ומשפיע על שינויים בטונוס השרירים החלקים (לא מפוספסים) של מערכת העיכול.
התפתחות בלוטת יותרת המוח
האונה הקדמית של בלוטת יותרת המוח מתפתחת מהאפיתל של הדופן הגבית של הפה בצורת גידול בצורת טבעת (כיס רתקה). בליטה אקטודרמלית זו צומחת לכיוון תחתית החדר השלישי העתידי. לכיוון זה, מהמשטח התחתון של שלפוחית המוח השנייה (התחתית העתידית של החדר השלישי), צומח תהליך, שממנו מתפתחים הפקעת האפורה של האונה האחורית של בלוטת יותרת המוח.
כלי דם ועצבים של בלוטת יותרת המוח
עורקי התרדמה הפנימיים וכלי הדם של מעגל העורקים של המוח, עורקי יותרת המוח העליונים והתחתונים מופנים לבלוטת יותרת המוח. עורקי יותרת המוח העליונים עוברים לגרעין האפור ולאינפונדיבולום של ההיפותלמוס, מתחברים זה לזה כאן ויוצרים נימים החודרים לרקמת המוח - רשת ההמוקפילרית הראשונית. מהלולאות הארוכות והקצרות של רשת זו נוצרים ורידי הפורטל, המופנים לאונה הקדמית של בלוטת יותרת המוח. בפרנכימה של האונה הקדמית של בלוטת יותרת המוח, ורידים אלה מתפרקים לנימים סינוסואידליים רחבים, ויוצרים את רשת ההמוקפילרית המשנית. האונה האחורית של בלוטת יותרת המוח מסופקת עם דם בעיקר על ידי עורק יותרת המוח התחתון. בין עורקי יותרת המוח העליונים והתחתונים יש אנסטומוזות עורקיות ארוכות. זרימת הדם הוורידי מרשת ההמוקפילרית המשנית מתבצעת דרך מערכת ורידים הזורמים לסינוסים המערות והבין-מערבות של הדורה מאטר של המוח.
סיבים סימפתטיים הנכנסים לאיבר יחד עם העורקים משתתפים בעצבוב של בלוטת יותרת המוח. סיבי עצב סימפתטיים פוסט-גנגליוניים יוצאים ממקלעת עורק התרדמה הפנימי. בנוסף, באונה האחורית של בלוטת יותרת המוח נמצאים קצוות רבים של תהליכים של תאי הפרשה עצבית הממוקמים בגרעיני ההיפותלמוס.
מאפיינים הקשורים לגיל של בלוטת יותרת המוח
המשקל הממוצע של בלוטת יותרת המוח אצל יילודים מגיע ל-0.12 גרם. משקל האיבר מכפיל את עצמו עד גיל 10 ומשולש עד גיל 15. עד גיל 20, משקל בלוטת יותרת המוח מגיע למקסימום (530-560 מ"ג) ונשאר כמעט ללא שינוי בתקופות גיל עוקבות. לאחר 60 שנה, נצפית ירידה קלה במשקל בלוטה אנדוקרינית זו.
 [ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ], [ 13 ], [ 14 ], [ 15 ]
[ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ], [ 13 ], [ 14 ], [ 15 ]
הורמוני יותרת המוח
אחדות הוויסות העצבים וההורמונלי בגוף מובטחת על ידי הקשר האנטומי והתפקודי ההדוק של בלוטת יותרת המוח וההיפותלמוס. קומפלקס זה קובע את מצב ותפקוד המערכת האנדוקרינית כולה.
בלוטת יותרת המוח העיקרית המייצרת מספר הורמונים פפטידיים המווסתים ישירות את תפקוד הבלוטות ההיקפיות היא בלוטת יותרת המוח. זוהי תצורה דמוית שעועית אדמדמה-אפורה המכוסה בקפסולה סיבית במשקל 0.5-0.6 גרם. היא משתנה מעט בהתאם למין ולגיל של האדם. החלוקה המקובלת של בלוטת יותרת המוח לשתי אונות הנבדלות בהתפתחותן, במבנהן ובתפקודן נותרה: האונה הדיסטלית הקדמית - אדנוהיפופיזה והאחורית - נוירוהיפופיזה. הראשונה מהווה כ-70% מהמסה הכוללת של הבלוטה ומחולקת באופן קונבנציונלי לחלקים הדיסטליים, התוך-קרקעיים והביניים, השנייה - לחלק האחורי, או האונה, ולגבעול יותרת המוח. הבלוטה ממוקמת בגומה של יותרת המוח של הסלה טורצ'יקה של עצם הספנואיד ומחוברת למוח דרך הגבעול. החלק העליון של האונה הקדמית מכוסה על ידי כיאזמה של הראייה ודרכי הראייה. אספקת הדם לבלוטת יותרת המוח שופעת מאוד ומתבצעת על ידי ענפים של עורק התרדמה הפנימי (עורקי יותרת המוח העליונים והתחתונים), כמו גם על ידי ענפים של מעגל העורקים של המוח. עורקי יותרת המוח העליונים משתתפים באספקת הדם לאדנוהיפופיזה, והתחתונים - הנוירוהיפופיזה, במגע עם הקצוות הנוירו-פרשיים של האקסונים של גרעיני התאים הגדולים של ההיפותלמוס. הראשונים נכנסים לקצה החציוני של ההיפותלמוס, שם הם מתפזרים לרשת נימים (מקלעת הנימים הראשונית). נימים אלה (איתם באים במגע טרמינלי האקסונים של תאי הנוירו-פרשיה הקטנים של ההיפותלמוס הבינוני-בסיסי) מתאספים לוורידים פורטליים היורדים לאורך גבעול יותרת המוח אל פרנכימה של האדנוהיפופיזה, שם הם מתחלקים שוב לרשת של נימים סינוסואידליים (מקלעת הנימים המשנית). לפיכך, הדם, לאחר שעבר בעבר דרך הגובה החציוני של ההיפותלמוס, שם הוא מועשר בהורמונים אדנוהיפופיזוטרופיים של ההיפותלמוס (הורמונים משחררים), נכנס לאדנוהיפופיזה.
זרימת הדם הרווי בהורמונים אדנוהיפופיזה מנימים רבים של מקלעת המשני מתבצעת דרך מערכת הוורידים, אשר בתורם זורמים לסינוסים הוורידיים של הדורה מאטר ומשם לזרם הדם הכללי. לפיכך, מערכת הפורטל של בלוטת יותרת המוח עם כיוון יורד של זרימת דם מההיפותלמוס היא מרכיב מורפופונקציונלי במנגנון המורכב של בקרה נוירוהומורלית של התפקודים הטרופיים של אדנוהיפופיזה.
בלוטת יותרת המוח (היפופיזה) מעוצבבת על ידי סיבים סימפתטיים העוקבים אחר עורקי יותרת המוח. מקורם בסיבים פוסט-גנגליוניים העוברים דרך מקלעת התרדמה הפנימית (internal carotid plexus), המחוברים לגנגליונים הצוואריים העליונים (superior cervical ganglia). אין עצבוב ישיר של האדנוהיפופיזה מההיפותלמוס. האונה האחורית מקבלת סיבי עצב מהגרעינים הנוירו-פרשניים של ההיפותלמוס.
האדנוהיפופיזה היא תצורה מורכבת מאוד בארכיטקטורה ההיסטולוגית שלה. יש לה שני סוגים של תאים בלוטיים - כרומופוביים וכרומופיליים. האחרונים מתחלקים בתורם לאצידופיליים ובזופיליים (תיאור היסטולוגי מפורט של בלוטת יותרת המוח ניתן בסעיף המתאים במדריך). עם זאת, יש לציין כי ההורמונים המיוצרים על ידי תאי הבלוטה המרכיבים את הפרנכימה של האדנוהיפופיזה, בשל גיוון האחרונים, שונים במידה מסוימת באופיים הכימי, והמבנה העדין של התאים המפרישים חייב להתאים לתכונות הביוסינתזה של כל אחד מהם. אך לעיתים ניתן לראות באדנוהיפופיזה צורות מעבר של תאים בלוטיים המסוגלים לייצר מספר הורמונים. ישנן עדויות לכך שסוג תאי הבלוטה של האדנוהיפופיזה לא תמיד נקבע גנטית.
מתחת לסרעפת של הסלע הטורצ'יקה נמצא החלק דמוי המשפך של האונה הקדמית. הוא חובק את גבעול בלוטת יותרת המוח, ונוגע עם הפקעת האפורה. חלק זה של האדנוהיפופיזה מאופיין בנוכחות תאי אפיתל ואספקת דם בשפע. הוא גם פעיל מבחינה הורמונלית.
החלק הביניים (האמצעי) של בלוטת יותרת המוח מורכב מכמה שכבות של תאים בזופיליים גדולים הפעילים בהפרשה.
בלוטת יותרת המוח מבצעת מגוון פונקציות באמצעות ההורמונים שלה. האונה הקדמית שלה מייצרת הורמונים אדרנוקורטיקוטרופיים (ACTH), הורמונים מגרים בלוטת התריס (TSH), הורמונים מגרים זקיק (FSH), הורמונים מחלמניים (LH), הורמונים ליפוטרופיים, כמו גם הורמון גדילה סומטוטרופי (STO) ופרולקטין. באונה האמצעית מסונתז הורמון מגרה מלנוציטים (MSH), ובאונה האחורית מצטברים וזופרסין ואוקסיטוצין.
ACTH
הורמוני יותרת המוח הם קבוצה של הורמוני חלבון, פפטיד וגליקופרוטאינים. מבין ההורמונים של בלוטת יותרת המוח הקדמית, ACTH נחקר בצורה הטובה ביותר. הוא מיוצר על ידי תאים בזופיליים. תפקידו הפיזיולוגי העיקרי הוא לעורר את הביוסינתזה וההפרשה של הורמוני סטרואידים על ידי קליפת האדרנל. ACTH מפגין גם פעילות מגרה מלנוציטים וליפוטרופית. בשנת 1953 הוא בודד בצורתו הטהורה. מאוחר יותר נקבע המבנה הכימי שלו, המורכב מ-39 שיירי חומצות אמינו בבני אדם ובמספר יונקים. ל-ACTH אין ספציפיות למין. כיום, בוצעה סינתזה כימית של ההורמון עצמו ושל שברים שונים של המולקולה שלו, פעילים יותר מהורמונים טבעיים. מבנה ההורמון מורכב משני חלקים של שרשרת הפפטידים, אחד מהם מבטיח את הגילוי והקישור של ACTH לקולטן, והשני מספק השפעה ביולוגית. הוא ככל הנראה נקשר לקולטן ACTH באמצעות אינטראקציה של מטענים חשמליים של ההורמון והקולטן. תפקיד האפקטור הביולוגי של ACTH מתבצע על ידי שבר של מולקולת 4-10 (Met-Glu-His-Phen-Arg-Tri-Tri).
הפעילות המגרה מלנוציטים של ACTH נובעת מנוכחותו במולקולה של האזור ה-N-טרמינלי, המורכב מ-13 שיירי חומצות אמינו וחוזר על המבנה של הורמון אלפא-מגרה מלנוציטים. אזור זה מכיל גם הפטאפפטיד הקיים בהורמוני בלוטת יותרת המוח אחרים ובעל פעילויות אדרנוקורטיקוטרופיות, מגרה מלנוציטים וליפוטרופיות מסוימות.
הרגע המרכזי בפעולת ACTH נחשב להפעלת האנזים חלבון קינאז בציטופלזמה בהשתתפות cAMP. חלבון קינאז זרחני מפעיל את האנזים אסטראז, אשר ממיר אסטרים של כולסטרול לחומר חופשי בטיפות שומן. החלבון המסונתז בציטופלזמה כתוצאה מזרחון הריבוזומים מגרה את קשירת הכולסטרול החופשי לציטוכרום P-450 ואת העברתו מטיפות השומנים למיטוכונדריה, שם נמצאים כל האנזימים המבטיחים את המרת הכולסטרול לקורטיקוסטרואידים.
[ 16 ], [ 17 ], [ 18 ], [ 19 ], [ 20 ], [ 21 ], [ 22 ]
הורמון מגרה בלוטת התריס
TSH - תירוטרופין - המווסת העיקרי של התפתחות ותפקוד בלוטת התריס, תהליכי הסינתזה וההפרשה של הורמוני בלוטת התריס. חלבון מורכב זה - גליקופרוטאין - מורכב מתת-יחידות אלפא ובטא. מבנה תת-היחידה הראשונה תואם לתת-היחידה אלפא של הורמון הלוטאין. יתר על כן, הוא תואם במידה רבה במינים שונים של בעלי חיים. רצף שיירי חומצות האמינו בתת-היחידה בטא של TSH אנושי פוענח והוא מורכב מ-119 שיירי חומצות אמינו. ניתן לציין שתת-היחידות בטא של TSH אנושי ובקר דומות במובנים רבים. התכונות הביולוגיות ואופי הפעילות הביולוגית של הורמוני גליקופרוטאין נקבעים על ידי תת-היחידה בטא. היא גם מבטיחה את האינטראקציה של ההורמון עם קולטנים באיברי מטרה שונים. עם זאת, תת-היחידה בטא ברוב בעלי החיים מציגה פעילות ספציפית רק לאחר שהיא מתחברת לתת-היחידה אלפא, הפועלת כמעין מפעיל של ההורמון. האחרון, בסבירות שווה, גורם לפעילויות לוטאין, מגרה זקיק ותירוטרופיות הנקבעות על ידי תכונות תת-היחידה בטא. הדמיון שנמצא מאפשר לנו להסיק כי הורמונים אלה נוצרו בתהליך האבולוציה מקודם משותף אחד, תת-היחידה בטא קובעת גם את התכונות החיסוניות של ההורמונים. קיימת הנחה שתת-היחידה אלפא מגינה על תת-היחידה בטא מפני פעולת אנזימים פרוטאוליטיים, וגם מקלה על הובלתה מבלוטת יותרת המוח לאיברי המטרה ההיקפיים.
הורמונים גונדוטרופיים
גונדוטרופינים קיימים בגוף בצורת LH ו-FSH. המטרה התפקודית של הורמונים אלה מצטמצמת בדרך כלל להבטחת תהליכי רבייה אצל אנשים משני המינים. הם, כמו TSH, הם חלבונים מורכבים - גליקופרוטאינים. FSH גורם להבשלת זקיקים בשחלות של נשים וממריץ יצירת זרע אצל גברים. LH גורם לקרע זקיק אצל נשים עם היווצרות הגופיף הצהוב וממריץ את הפרשת האסטרוגנים והפרוגסטרון. אצל גברים, אותו הורמון מאיץ את התפתחות הרקמה הבין-סטיציאלית ואת הפרשת האנדרוגנים. השפעות הגונדוטרופינים תלויות זו בזו ומתרחשות באופן סינכרוני.
הדינמיקה של הפרשת גונדוטרופינים אצל נשים משתנה במהלך המחזור החודשי ונחקרה בפירוט מספק. בשלב הטרום-ביוצי (הזקיקים) של המחזור, תכולת ה-LH נמצאת ברמה נמוכה למדי, בעוד ש-FSH עולה. ככל שהזקיק מתבגר, הפרשת האסטרדיול עולה, מה שמקדם ייצור מוגבר של גונדוטרופינים על ידי בלוטת יותרת המוח ואת התרחשותם של מחזורי LH ו-FSH כאחד, כלומר סטרואידים מיניים מגרים את הפרשת הגונדוטרופינים.
נכון לעכשיו, נקבע מבנה ה-LH. כמו TSH, הוא מורכב משתי תת-יחידות: a ו-b. מבנה תת-היחידה האלפא של LH במינים שונים של בעלי חיים חופף במידה רבה, הוא תואם את מבנה תת-היחידה האלפא של TSH.
מבנה תת-היחידה בטא של LH שונה באופן משמעותי ממבנה תת-היחידה בטא של TSH, למרות שיש לה ארבעה מקטעים זהים של שרשרת הפפטידים המורכבים מ-4-5 שיירי חומצות אמינו. ב-TSH, הם ממוקמים במיקומים 27-31, 51-54, 65-68 ו-78-83. מכיוון שתת-היחידה בטא של LH ו-TSH קובעת את הפעילות הביולוגית הספציפית של ההורמונים, ניתן להניח שמקטעים הומולוגיים במבנה של LH ו-TSH אמורים להבטיח את הקשר של תת-היחידות בטא לתת-היחידה אלפא, ומקטעים הנבדלים במבנה אמורים להיות אחראים לספציפיות של הפעילות הביולוגית של ההורמונים.
LH טבעי יציב מאוד לפעולת אנזימים פרוטאוליטיים, אך תת-היחידה בטא מתפרקת במהירות על ידי כימוטריפסין, ותת-היחידה a קשה לעבור הידרוליזה על ידי האנזים, כלומר היא ממלאת תפקיד מגן, המונע מכימוטריפסין גישה לקשרים פפטידיים.
באשר למבנה הכימי של FSH, החוקרים טרם השיגו תוצאות חד משמעיות. כמו LH, FSH מורכב משתי תת-יחידות, אך תת-היחידה בטא של FSH שונה מתת-היחידה בטא של LH.
פרולקטין
הורמון נוסף, פרולקטין (הורמון לקטוגני), ממלא תפקיד פעיל בתהליכי רבייה. התכונות הפיזיולוגיות העיקריות של פרולקטין אצל יונקים מתבטאות בצורה של גירוי התפתחות בלוטות החלב וההנקה, גדילת בלוטות החלב והאיברים הפנימיים. הוא מקדם את ביטוי השפעת הסטרואידים על מאפיינים מיניים משניים אצל זכרים, מגרה את פעילות ההפרשה של הגופיף הצהוב בעכברים וחולדות, ומשתתף בוויסות חילוף החומרים של שומנים. תשומת לב רבה הוקדשה בשנים האחרונות לפרולקטין כווסת התנהגות אימהית; רב-תפקודיות זו מוסברת על ידי התפתחותו האבולוציונית. זהו אחד מהורמוני יותרת המוח העתיקים והוא נמצא אפילו אצל דו-חיים. כיום, מבנה הפרולקטין אצל כמה מיני יונקים פוענח לחלוטין. עם זאת, עד לאחרונה, מדענים הביעו ספקות לגבי קיומו של הורמון כזה בבני אדם. רבים האמינו שתפקידו מבוצע על ידי הורמון גדילה. כעת הושגו ראיות משכנעות לנוכחות פרולקטין בבני אדם ומבנהו פוענח חלקית. קולטני פרולקטין נקשרים באופן פעיל להורמון גדילה ולקטוגן השליה, דבר המצביע על מנגנון פעולה יחיד של שלושת ההורמונים.
סומטוטרופין
להורמון הגדילה, סומטוטרופין, יש ספקטרום פעולה רחב אף יותר מזה של פרולקטין. כמו פרולקטין, הוא מיוצר על ידי תאים אצידופיליים של אדנוהיפופיזה. סומטוטרופין מגרה גדילה שלדית, מפעיל ביוסינתזה של חלבונים, בעל השפעה על גיוס שומנים ומקדם עלייה בגודל הגוף. בנוסף, הוא מתאם תהליכים מטבוליים.
השתתפותו של ההורמון באחרון מאושרת על ידי עלייה חדה בהפרשתו על ידי בלוטת יותרת המוח, למשל, כאשר רמת הסוכר בדם יורדת.
המבנה הכימי של הורמון אנושי זה נקבע במלואו כעת - 191 שיירי חומצות אמינו. המבנה הראשוני שלו דומה לזה של סומטוממוטרופין כוריוני או לקטוגן שליה. נתונים אלה מצביעים על קרבה אבולוציונית משמעותית בין שני ההורמונים, אם כי הם מפגינים הבדלים בפעילות הביולוגית.
יש להדגיש את הספציפיות הגבוהה למין של ההורמון המדובר - לדוגמה, הורמון גדילה אנושי שמקורו מן החי אינו פעיל בבני אדם. זה מוסבר הן על ידי התגובה בין קולטני הורמון גדילה אנושי של בני אדם ובעלי חיים, והן על ידי מבנה ההורמון עצמו. כיום, נערכים מחקרים לזיהוי מרכזים פעילים במבנה המורכב של הורמון גדילה המציגים פעילות ביולוגית. נחקרים שברים בודדים של המולקולה המציגים תכונות אחרות. לדוגמה, לאחר הידרוליזה של הורמון גדילה אנושי על ידי פפסין, בודד פפטיד המורכב מ-14 שיירי חומצות אמינו ומתאים לחלק המולקולה 31-44. לא הייתה לו השפעה על גדילה, אך עלה משמעותית על ההורמון הטבעי בפעילות ליפוטרופית. הורמון גדילה אנושי, בניגוד להורמון דומה של בעלי חיים, בעל פעילות לקטוגנית משמעותית.
האדנוהיפופיזה מסנתזת חומרים פפטידיים וחלבוניים רבים בעלי השפעה על גיוס שומן, ולהורמונים הטרופיים של בלוטת יותרת המוח - ACTH, STH, TSH ואחרים - יש השפעה ליפוטרופית. בשנים האחרונות, הורמונים בטא ו-β-ליפוטרופיים (LPG) הודגשו במיוחד. התכונות הביולוגיות של בטא-LPG נחקרו בפירוט רב; בנוסף לפעילות ליפוטרופית, יש לו גם השפעה מגרה מלנוציטים, מגרה קורטיקוטרופין והיפוקלצמית, והוא גם מייצר השפעה דמוית אינסולין.
כיום, פוענח המבנה הראשוני של LPG בכבשים (90 שיירי חומצות אמינו), הורמונים ליפוטרופיים של חזירים ובקר. להורמון זה יש ספציפיות למין, אם כי מבנה האזור המרכזי של בטא-LPG זהה במינים שונים. הוא קובע את התכונות הביולוגיות של ההורמון. אחד השברים של אזור זה נמצא במבנה של אלפא-MSH, בטא-MSH, ACTH ובטא-LPG. ההנחה היא שהורמונים אלה נבעו מאותו תוצר קדם בתהליך האבולוציה. ל-γ-LPG יש פעילות ליפוטרופית חלשה יותר מאשר בטא-LPG.
הורמון מגרה מלנוציטים
הורמון זה, המסונתז באונה האמצעית של בלוטת יותרת המוח, מגרה את הביוסינתזה של פיגמנט העור מלנין בתפקודו הביולוגי, מקדם עלייה בגודל ובמספר תאי הפיגמנט (מלנוציטים) בעורם של דו-חיים. תכונות אלו של MSH משמשות בבדיקות ביולוגיות של ההורמון. ישנם שני סוגים של ההורמון: אלפא- ובטא-MSH. הוכח כי לאלפא-MSH אין ספציפיות למין ויש לו את אותו המבנה הכימי בכל היונקים. המולקולה שלו היא שרשרת פפטידית המורכבת מ-13 שיירי חומצות אמינו. לעומת זאת, לבטא-MSH יש ספציפיות למין, ומבנהו שונה בבעלי חיים שונים. ברוב היונקים, מולקולת בטא-MSH מורכבת מ-18 שיירי חומצות אמינו, ורק בבני אדם היא מורחבת מקצה חומצת האמינו בארבע שיירי חומצות אמינו. יש לציין כי לאלפא-MSH יש פעילות אדרנוקורטיקוטרופית מסוימת, והשפעתו על התנהגותם של בעלי חיים ובני אדם הוכחה כעת.
אוקסיטוצין ווזופרסין
וזופרסין ואוקסיטוצין, המסונתזים בהיפותלמוס, מצטברים באונה האחורית של בלוטת יותרת המוח: וזופרסין בנוירונים של הגרעין הסופראופטי, ואוקסיטוצין בגרעין הפרה-חדרי. לאחר מכן הם מועברים לבלוטת יותרת המוח. יש להדגיש כי ההורמון המבשר של הורמון וזופרסין מסונתז תחילה בהיפותלמוס. במקביל, שם מיוצרים החלבונים נוירופיזין מסוג 1 ו-2. הראשון נקשר לאוקסיטוצין, והשני נקשר לוזופרסין. קומפלקסים אלה נודדים בצורת גרגירים נוירו-פרשתיים בציטופלזמה לאורך האקסון ומגיעים לאונה האחורית של בלוטת יותרת המוח, שם סיבי העצב מסתיימים בדופן כלי הדם ותוכן הגרגירים נכנס לדם. וזופרסין ואוקסיטוצין הם הורמוני יותרת המוח הראשונים עם רצף חומצות אמינו מבוסס לחלוטין. במבנה הכימי שלהם, הם לא-אפפטידים עם גשר דיסולפידי אחד.
ההורמונים הנדונים מייצרים מגוון השפעות ביולוגיות: הם מגרים את הובלת המים והמלחים דרך הקרומים, בעלי השפעה של לחץ דם, משפרים את התכווצויות השרירים החלקים של הרחם במהלך הלידה ומגבירים את הפרשת בלוטות החלב. יש לציין כי לוזופרסין פעילות אנטי-דיורטית גבוהה יותר מאשר לאוקסיטוצין, בעוד שלאחרון יש השפעה חזקה יותר על הרחם ועל בלוטת החלב. הרגולטור העיקרי של הפרשת וזופרסין הוא צריכת מים; בצינורות הכליות, הוא נקשר לקולטנים בקרומים הציטופלזמיים ולאחר מכן מפעיל את האנזים אדנילט ציקלאז בהם. חלקים שונים של המולקולה אחראים על קשירת ההורמון לקולטן ועל ההשפעה הביולוגית.
בלוטת יותרת המוח, המחוברת דרך ההיפותלמוס עם מערכת העצבים כולה, מאחדת את המערכת האנדוקרינית לשלם פונקציונלי, ומשתתפת בהבטחת הקביעות של הסביבה הפנימית של הגוף (הומאוסטזיס). בתוך המערכת האנדוקרינית, מתבצע ויסות הומאוסטטי על סמך עקרון המשוב בין האונה הקדמית של בלוטת יותרת המוח לבלוטות "המטרה" (בלוטת התריס, קליפת האדרנל, בלוטות המין). עודף של הורמון המיוצר על ידי בלוטת "המטרה" מעכב, וחסר בו מגרה את ההפרשה והשחרור של ההורמון הטרופי המתאים. ההיפותלמוס נכלל במערכת המשוב. בה נמצאים אזורי הקולטנים הרגישים להורמונים של בלוטות "המטרה". על ידי קישור ספציפי להורמונים במחזור הדם ושינוי התגובה בהתאם לריכוז ההורמונים, קולטני ההיפותלמוס מעבירים את השפעתם למרכזי ההיפותלמוס המתאימים, אשר מתאמים את עבודת האדנוהיפופיזה, ומשחררים הורמונים אדנוהיפופיזוטרופיים של ההיפותלמוס. לפיכך, יש להתייחס להיפותלמוס כמוח נוירואנדוקריני.